La nutrición como terapia antienvejecimiento.
Desde hace mucho tiempo se ha relacionado la nutrición con el envejecimiento. Esta relación se ha centrado, básicamente, en los estudios de restricción calórica y en la suplementación con sustancias antioxidantes. Más recientemente se han realizado estudios encaminados a estudiar el papel de las grasas en las dieta, desde el punto de vista del estrés oxidativo como terapia antienvejecimiento.
Restricción calórica.
Como se ha descrito antes, la restricción calórica aumenta la vida media en un amplio rango de especies y disminuye la velocidad con que aparecen determinadas enfermedades relacionadas con la edad. Este efecto se lleva a cabo a través de una reducción en el estrés oxidativo. Esto se sustenta, entre otras pruebas, en la observación de que ratones calóricamente restringidos generan un menor estrés oxidativo que sus homólogos alimentados ad libitum, produciendo además un menor índice de oxidación de lípidos, proteínas, y ADN.
La restricción calórica, además previene mucho de los cambios que se produce en relación a la expresión génica durante el envejecimiento, ente los que se incluyen la elevación en la expresión de las proteínas de shock término y la atenuación de la expresión de la proteína inducida por estrés Hsp70.
La restricción calórica podría ser una potente arma terapéutica para lucha contra el envejecimiento, ya que en un principio cumple con los requisitos exigibles de efectividad frente a la reducción del estrés oxidativo y el retraso del envejecimiento, así como contra las enfermedades asociadas a éste. No obstante, la posible aplicación de la restricción calórica como terapia antienvejecimiento en la población humana acarrea tales dificultades éticas y de tipo práctico que hacen prácticamente inviable su puesta en marcha.

Antioxidantes.
Como se ha puesto de manifiesto en los apartados anteriores, el estrés oxidativo desempeña un papel muy significativo en el proceso global de envejecimiento y, por lo tanto, la suplementación con antioxidantes podría ser de utilidad como posible terapia antienvejecimiento. Entre los primeros estudios cabe destacar los de Miquel y Ecónomos en relación a la capacidad del carbosilato de tiazolidina de aumentar la vitalidad y programar la vida media en ratones.
Posteriormente, Furukawa y cols. Mostraron el papel protector de la administración oral del glutatión frente al declive de la función inmunológica asociada al envejecimiento. Muchos otros antioxidantes han sido probados en relación al envejecimiento, con resultados más o menos positivos.
Entre dichos antioxidantes cabe destacar las vitaminas E y C, la coenzima Q, extractos herbales ricos en flavonoides y polifenoles, entre otros. Si bien los resultados obtenidos con estos antioxidantes han sido exitosos atenuación del estrés oxidativo mediado por la edad o por enfermedades asociadas a éstas, han tenido poco o ningún éxito en relación al aumento de la longevidad.
Tal vez para tener un mayor éxito con la terapia basada en antioxidantes se debería profundizar en el conocimiento de las propiedades farmacológicas de las sustancias empleadas, sobre todo en lo concerniente a la absorción, la distribución tisular y el metabolismo de estas. Además, no debe olvidarse el papel que las ROS tienen en la señalización celular, de modo que la dosis de antioxidante debe ser muy bien ajustada para evitar cambios en el estado redox que podrían alterar la función celular. Los problemas anteriores estas siendo solucionados, en parte, mediante el uso de una nueva generación de sustancias antioxidantes sintéticas, mimñeticos de la superóxido dismultasa y la catalasa. Estas sustancias están siendo ensayadas con cierto éxito, habiéndose mostrado efectivas en el aumento de la longevidad en ratones y C. elegans.
Dentro del grupo de polifenoles merece especial atención el resveratrol. Esta sustancia ha demostrado extender la vida media en diversos organismos, así como ejercer un efecto positivo en múltiples enfermedades, como el cáncer, enfermedad cardiovascular, Alzheimer, enfermedades metabólicas, entre otras. Su principal mecanismo de acción es mediante la activación de la SIRT1 (miembro de la familia denominada sirtuinas), mimetizando así el efecto de restricción calórica sobre el envejecimiento. Estudios recientes muestran otra diana directa del reveratrol, la PI3K (fosfatidilinositol-3-quinasa), representándose asi otra via central de control de la vida máxima independiente de situinas.

Ácidos grasos de la dieta.
El tipo de grasa de la dieta condiciona de manera importante numerosos parámetros bioquímicos en la membrana mitocondrial. La importancia del tipo de ácidos grasos de la dieta reside en el hecho de que la membrana mitocondrial (y, en general, todas las membranas biológicas) es capaz de adaptar la composición de sus fosfolípidos a la grasa ingerida de forma mayoritaria. De este modo, si un individuo ingiere mayoritariamente grasa de origen animal, sus membranas serán más ricas en ácidos grasos saturados que las de otro individuo cuya fuente grasa mayoritaria sea de origen vegetal.
Por otro lado, se ha descrito de forma contundente cómo se producen las adaptaciones del sistema de transporte electrónico mitocondrial en relación al tipo de grasa en la dieta, con mayor o menor repercusión sobre los diversos complejos del sistema. Además el estrés oxidativo está relacionado con la composición lipídica de las membranas biológicas, de modo que una fuente grasa poliinsaturada (p. ej., aceite de girasol) generará membranas más susceptibles al daño oxidativo que una fuente saturada (grasa animal) o monoinsaturada (aceite de oliva), lo cual ha sido ampliamente demostrado en numerosas situaciones fisiológicas y patológicas, empleando numerosos modelos animales y humanos.
Los resultados obtenidos en este campo apuntan las siguientes conclusiones: el envejecimiento, entendido como un proceso endógeno y progresivo, provoca a lo largo de la vida alteraciones en la mitocondria y sus componentes, como el mtDNA (alteraciones que tienen un elevado comportamiento oxidativo). Estas alteraciones deterioran la estructura y la función mitocondriales, y, dependiendo de la capacidad del tejido en concreto para reparar el daño o eliminar la célula alterada, la función tisularar se verá afectada en mayor o menor grado.
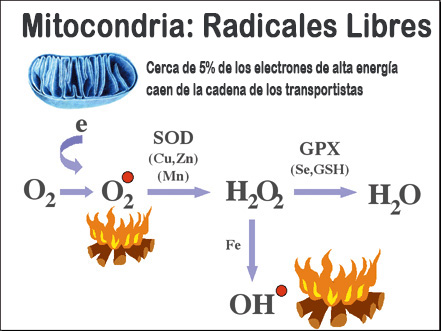
Así, los tejidos con capacidad regenerativa, como el hígado, parecen ser capaces de remediar el daño ocasionado, como lo siguiente la ausencia de pérdida de función mitocondrial en términos de actividad citocomo axidasa. Sin embargo, se produce una pérdida de función en los tejidos posmitóticos, como el musculo esquelético o el corazón, sin capacidad para reemplazar células y, probablemente, con un sistema de reparación de daño menos efectivo (existen diferencias entre el hígado y el corazón, en relación al sistema de reparación del mtDNA). Esta pérdida de función se refleja en el descenso brusco de la actividad citocromo oxidasa, lo que da lugar a un desacoplamiento de la CTEmt, con la consiguiente ineficacia bioenergética y el aumento en la producción de ROS.
Las mitocondrias de los tejidos posmitóticos tratan de atenuar la situación desfavorable mediante el aumento de otros componentes de la CTEmt, como el citocromo b, o por medio de aumentos en el grado de poliinsaturación probablemente para intentar aumentar la fluidez y la actividad de la citocromo oxidasa restante mediante la presencia de una cardiolipina más poliinsaturada. Sin embargo, ambas acciones generan un aumento mayor en la producción de radicales libres.
El papel de la grasa en la dieta en este mecanismo residiría, por lo tanto, en la construcción de un entorno más o menos susceptible para la generación y la programación de ROS, especialmente cuando, como consecuencia de los procesos como el envejecimiento, se producen fallos en la CTEmt.